運動反射と運動制御理論
(バイオメカニズム学会誌より)道免和久
要 旨
運動反射の観点から運動制御理論を概説した。代表的な運動制御理論として、反射を制御の中心としたサーボ仮説(γmodel)、緊張性伸張反射機構に対して筋長を規定する中枢指令λを仮定したλmodel、反射を神経系のハードウェアの一部としたフィードフォワード系中心の内部モデル制御理論、求心情報自体を必要としないαmodelがある。反射中心に制御するγmodelは現在信じられておらず、反射回路を含まないαmodelも特殊な制御とされる。λmodelは部分的に否定されているが、N型仮想軌道によるλmodelについて決着はついていない。内部モデルによる制御理論がどのように各レベルの反射を包括していくのか、今後の発展が期待される。
1.はじめに
人間の随意運動は多様性に満ち、一見ステレオタイプにみえる運動でも背後には冗長な自由度をもつ筋骨格系を動かす複雑かつ合目的的なシステム、すなわち脳が存在している。複雑な制御システムに比較すれば、一定のパターンを示す運動反射は単純な部品の一つと考えるべきであろう。もちろん、一つの中枢神経細胞を介する単シナプス反射でさえも、本特集の解説(小宮山,1999)に記述されているように、その入出力関係には複雑な要素がからんでおり、運動制御にかかわる多くの問題を含んでいることに異論はない。しかし、反射だけで随意運動制御のすべてを説明しようとすることは明らかに誤りである。逆に、反射を脳幹や脊髄などに組み込まれた神経系のハードウエアとして解明し、反射を脳がどのように制御しているか、随意運動制御の中で反射がどのような役割を演じているかを理解する方が運動の本質に近いはずである。特に臨床系の分野においてこの点について誤解が多いので最初に強調しておきたい。
本総説では主要な運動制御理論と反射との関連を中心に概説する。
2.ReactionとAction
運動制御と反射をめぐって歴史的に象徴的な出来事を紹介しよう。随意運動における冗長性の問題などを50年以上前に指摘し、今日の運動制御研究の基礎を開いたのはロシアのNicholai A Bernsteinである(Bernstein NA, 1967)。当時、Pavlovの条件反射理論が趨勢であり、動物はすべて外界からの刺激に反応して動く、あるは、行動は環境からの刺激によって規定される、という理論や人間観が広く受け入れられていた。Pavlovは、脳を各種反射の相互作用に基づいて受動的に反応するだけの臓器ととらえていた。Bernsteinはこの考え方に異論を唱え、能動的に環境を探索することがより動物の脳の本質に近いと主張した(Ferigenberg,1996)。そして、"re-action"を研究する伝統的な生理学ではなく、"action"、すなわち随意運動の研究を本格的に開始し、今日の運動制御研究の基礎を確立したことから、神経科学の改革者と言われるようになった。ところが、彼の"Physiology of action"という考えは、"西側"の科学の受け売りと学会で批判されただけでなく、当時の政府によって危険思想ととらえられ、プラウダ誌で名指しで批判されたために、彼は職を失ってしまったという不幸なエピソードがある。そして近年になってようやくBernsteinは世界的に再認識されるようになった。このように、反射を含む"reaction"の研究は、随意運動あるいは行動"action"の研究を排除し、対立してきたという残念な歴史がある。
現在でも依然として、一定の入力に対して一定の再現性を有する反応が得られる反射は、データとして信頼され、とくにわが国では"reaction"の研究が重視されているようである。コンピュータの発達によって波形解析などデータ処理が容易になってきた現在では、単一ニューロン活動の記録、精密3次元動作解析、非侵襲脳機能測定法、筋電図などを利用して、国際的に運動制御に関する研究業績が急増している。データの「再現性」の殻だけに閉じこもって"reaction"だけをみていては、随意運動制御の本質が明らかになることはないであろう。随意運動制御は反射のような一定のデータではなく、複雑なシステムを反映して、ある程度再現性があるが、ある程度多様なデータになるからであり、このようなデータから、脳の情報処理などの機能を理論立てていく作業が今後ますます重要になると考えられる。この作業の中で脳の運動制御における反射の役割も同時に明らかになっていくものと考える。
3.運動制御理論における運動反射の位置
運動制御理論において、運動反射の概念を包括した理論は必ずしも多くはない。随意運動中、伸張反射は運動の妨げになるので、フィードバック系は抑制されていることから、運動中は反射の役割は小さいとする考えがある。さらに、後述する有名なBizziの研究(Bizzi,1982)のように、感覚情報がなくても随意運動制御が可能であるという説がある。一方で、運動制御においてフィードバックループを積極的に利用している説もある。
運動反射の観点から運動制御理論をまとめると、1)反射は随意運動の主たる制御機構であるという説(γ model)、2)随意運動は反射のパラメータをうまく調節して制御されるという説(λmodel)、3)反射は随意運動の補助にすぎないという説(αmodel)、4)反射は随意運動の一部品であるという説(内部モデルによる制御)にわけられる。以下、各理論について解説する。
4.Mertonのサーボコントロール仮説(γmodel)
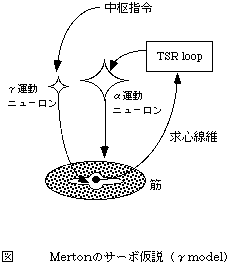
Merton(Merton PA 1953)は緊張性伸張反射tonic stretch reflexが主たる運動制御メカニズムと考え、サーボ仮説servo hypothesisを提唱した(図1)。伸張反射という神経生理学的な機構をはじめて制御の理論に取り入れた仮説である。γ運動ニューロンへの入力が筋長を制御する機構であることから、γmodelとも呼ばれる。
図1 Mertonのサーボ仮説(γ model)
運動指令はγ運動ニューロンに与えられる。情報の流れは、γ運動ニューロン活動→筋紡錐→緊張性伸張反射(tonic steretch reflex:TSR)→α運動ニューロン活動→筋張力の発生→運動、のようになっている。
初期には、同名筋に対するIa群線維を介した単シナプス反射をモデル化したが、後に、緊張性伸張反射に基づいて理論化した。γmodelにおいて緊張性伸張反射の機構は、筋肉が収縮によって短縮すれば筋紡錐の活動は低下する、というようにネガティヴ・フィードバックの機構として働く。完全なサーボ機構によって筋長が制御されているとすると、外力が加わっても筋長が変化しない、すなわち、無限大の利得をもつことになる。しかし、これまで緊張性伸張反射がそのような高い利得をもつことについては否定されている(Hoffer & Andreassen, 1981)。
運動制御理論に神経生理学的な知見を取り入れた点では、歴史的に重要であり、基本的な運動制御理論としての重要性は十分認めるべきであるが、反射とフィードバック機構中心の運動制御を固定観念にしてしまった理論でもある。
5.α-γ共賦活によるコントロール(α-γmodel)
γmodelでは、γ運動ニューロンへの入力が運動指令となるので、α運動ニューロンの活動は緊張性伸張反射のフィードバックループの分、運動の発現には遅れが生じる。このような理論的欠点を補う現象として、α運動ニューロンとγ運動ニューロンが同時に活動すること、すなわちα-γ共賦活現象がある(Granit, 1970)。この現象から、α運動ニューロンへのフィードフォワード運動指令とγ運動ニューロンへの長さ制御の運動指令の組み合わせによって、随意運動が発現するというα-γ運動ニューロンによる制御理論に発展した。しかしこれは、γmodelと同様に、基本的に緊張性伸張反射が高い利得で筋長を規定することが前提になっているので、制御理論として否定的である。
したがって、α-γ共賦活自体は、筋肉が収縮して短縮することによって筋紡錐の感受性が低下するのを防ぐ生理学的なメカニズムとしてとらえるべきである。
6.αmodel
脳がα運動ニューロンの活動を主に制御するという仮説である。Bizziら(Bizzi,1982, Bizzi, 1983)は、肘関節を一定の角度屈曲させる運動課題をサルに覚えさせ、そのサルの脊髄後根を切断(求心遮断:deafferentiation)しても、同じ運動課題が可能であったことから、緊張性伸張反射を介することなく、α運動ニューロンだけで随意運動が発現するという説を提唱した。すなわち、中枢の運動指令は、α運動ニューロンプールの活動レベルを変化させ、筋肉のバネ特性を変化させることにより、主動筋と拮抗筋のつり合い位置が変化して随意運動が発現する(図2)。
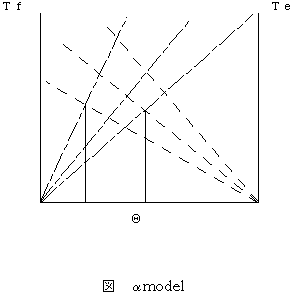
図2 α model
屈筋と伸筋のα運動ニューロンの活動性によって関節位置が制御される。屈筋張力Tfと伸筋張力Teとのつりあい位置が、関節の位置(Θ)となる。
このつり合い位置が描く軌道が、仮想軌道と呼ばれ、仮想軌道制御仮説とも呼ばれる。二つのばねがつり合う位置、すなわち平衡点が変化することが運動制御の本質と考えたことから、事項で説明するλmodelと混同されることがあるが、制御におけるフィードバックループの考え方が全く異なるため、本質的に別の理論と言ってよい。Latash(Latash, 1993)は、αmodelは求心遮断という特殊な状況下において中枢神経系が利用せざるをえなくなった特殊な運動制御であると指摘している。また、αmodelにおける仮想軌道は、実現軌道の鋳型のように近い形をしていることが前提となっているが、そのためには運動中の筋骨格系が高いスティフネスをもたなければならない。しかし、実際には運動中のスティフネスは小さく抑えれ、やわらかい制御が行われていることがわかっている(川人, 1996)。
7.λ model
中枢指令が制御するのは、緊張性伸張反射の閾値であり、筋長がこの閾値より長いときは筋肉が活動し、逆に短ければ筋活動は停止する。この仮説をλ model(Feldman, 1966, Feldman 1986, Latash 1993, Latash 1998)といい、λは緊張性伸張反射の閾値となる筋長をさす。この仮説はサーボ仮説で問題となった高い利得を仮定しない。すなわち、下行する中枢指令が一定のとき、平衡点のあつまりは長さ張力曲線上で一本の曲線を描き、フィードバックによる筋長調節の利得はこの曲線で規定される。これをinvariant characteristics(IC)と呼び、中枢指令はこのIC曲線の位置、つまり、緊張性伸張反射の閾値を変化させることによって随意運動を発現させる(図3)。Renshow抑制は、IC曲線の傾きを小さくし、スティフネスは減少する。また、相反性抑制が高まれば、IC曲線の傾きが大きくなり、スティフネスは増加する。また、主動筋と拮抗筋に対する運動指令として、位置制御の指令r(reciprocal )commandとスティフネス制御の指令c(coactivation) commandを想定している。主動筋のλをλago、拮抗筋のλをλantとするとき、rcommandは、(λag + λant)/2、c commandは (λag - λant)/2 であらわされる。中枢神経はこれら2種の指令で運動を制御していると仮定している。
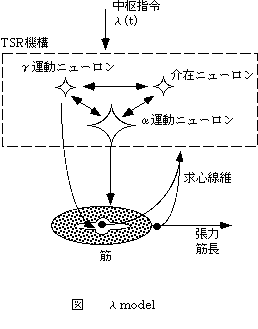
図3 λmodel
中枢指令は緊張性伸張反射TSRの機構全体に送られる。TSR機構はすべての末梢受容器の反射情報を受け取る。λmodelでは筋長、張力、筋活動、求心情報いずれもが制御において重要な役割をはたしている。
さらに、λの概念を 相動性の変化に拡張し、運動中の筋活動を理解することができる。運動中のλをλ*として、λ*=λ-μvと仮定する。ここで、μは定数、vは筋の短縮速度である。この関係式から、速度-長さ平面上に筋活動領域を定義できる。筋活動はこの領域の境界線(Ω直線)から遠ざかるほど、大きくなる。筋活動領域上にΩ直線に近い方から、小さな運動単位が配列されていると考えると、Hennemanのsize principleが理解できる。単関節の急速運動において、主動筋と拮抗筋に3相性の筋電パターンが出現することが知られているが、速度-長さ平面上で、3相性の筋電パターンが再現できる(図4)。
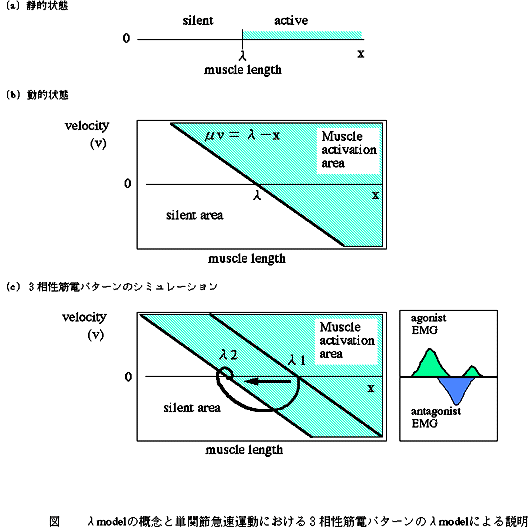
図4 λmodelの概念と単関節急速運動における3相性筋電パターンのλmodelによる説明
(a)静的状態:筋長xにおける運動ニューロンの活動条件はx-λ>0である。そして、活動する運動ニューロンの数はx-λに比例する。
(b)動的状態:筋肉がある一定のスピードで伸張しているとき、伸張反射の閾値は速度に比例して静的状態より小さくなる。即ち、λ*(dynamic λ)<λ (static λ)。 従って、λ* = λ-μdx/dt となる(μは定数)。この関係式から動的状態においてはx>λ-μdx/dtのとき運動ニューロンが活動するということになり、xとdx/dtの関係を表したグラフ上でこの関係を満たす領域 Muscle activation area を描くことができる。活動する運動ニューロンの数は静的状態と同様 x-λ*=x+μdx/dt-λに比例する。
(c)3相性筋電パターンのシミュレーション:λがλ1からλ2に変化した場合、図のような軌道をとってλ2に達する。Muscle activation area の範囲内で主動筋が活動する。完全にreciprocalなモデルを仮定すると、Muscle activation area の範囲外で拮抗筋が活動する。
Feldmanのλmodelにおいて、仮想軌道は実現軌道に近い形となっている。これは、Feldmanが高い粘性を仮定しているからであるが、Latashのλmodelでは低い粘性を仮定しているため、N型といわれるような仮想軌道になっている。仮想軌道が鋳型のように実際の軌道に先行するという点では、αmodelとFeldmanのλmodelは共通している。そして、このような単純な仮想軌道の計算には複雑な逆ダイナミクスの計算は脳内には必要ない、ということが、仮想軌道による制御を主張する研究者たちのもう一つの主張であった。しかし、Gomi ら(Gomi, 1996)が示したように、運動中の仮想軌道は、かなり複雑な形をしているため、仮想軌道制御については疑問視されつつある。Latashのλmodelについては、もともとN型の仮想軌道を計算で求めており、中枢神経がこのような仮想軌道のパターンを出力していると主張しているため(Latash , 1992, Domen 1999)、まだ決着はついていない。
λmodelは多くの批判にさらされながら、20年あまりにわたって主要な運動制御理論としての位置を保ってきた。この理論は、脊髄の反射を基本的な制御のメカニズムとしながらも、中枢神経がフィードフォワードの運動指令を出力することによって、複雑な随意運動が発現するとした点で、画期的であった。脳内でどのような計算が行われているか、というアプローチが主流となってきた現在、脳内の計算をブラックボックスとみなすλmodelはその歴史的役割を終えようとしている。しかし、仮想的緊張性伸張反射を仮定することで、従来の反射学における問題にも一応の回答を与えることができ、それを含みながら、随意運動全体を簡潔な理論である程度説明することができた点で、すぐれた理論といえる。
8.内部モデルによる運動制御
運動単位、反射、脊髄神経などの研究から、中枢神経における運動制御のメカニズムを解明しようとするボトムアップアプローチが伝統的な方法であった。逆に、神経系のハードウェアを十分に尊重しながら、脳における表現とアルゴリズムそして計算理論を明らかにするというトップダウンアプローチにより運動制御のメカニズムがわかってきた。このような方法論を計算論的神経科学(川人, 1996)という。
小脳皮質は層状の均一な構造をしており、学習が可能な神経回路ニューラルネットワークの集まりであると考えられている。運動中の学習によって小脳は、筋骨格系への入出力関係、つまり、運動指令とその結果として生じる軌道との関係の情報を蓄える。このような小脳内に保持される筋骨格系の情報は内部モデルと呼ばれ、運動指令から軌道を出力する神経回路を順モデル、逆に軌道に見合った運動指令を出力する神経回路を逆モデルと呼ぶ。
川人は、内部モデルの概念を含む運動制御理論、すなわちフィードバック誤差学習理論(Kawato, 1987)を提案している(図5)。まず連合野から運動野に目標軌道が送られ、運動野から運動指令が脊髄へ伝えられる。実現した運動についての感覚情報は、脊髄を上行し、大脳皮質レベルでのフィードバックループとして知られているトランスコーチカルループによって体性感覚野から運動野にフィードバックされる。このフィードバック回路でも運動は可能だが、フィードバック時間の遅れなどの理由で、速いなめらかな運動はできない。そこで、小脳-赤核系は、運動指令と軌道の関係を表現する内部モデルを学習によって形成し、速いスムーズな運動を可能にするとともに運動前に軌道誤差を予測して運動指令の修正を行うことを可能にする。目標軌道と実現軌道の間の誤差は、誤差信号(教師信号)として下オリーブ核から小脳に伝達され、小脳の内部モデルの修正が行われる。これは、運動制御と運動学習を統合した理論として知られている。多くの研究により内部モデルによる制御理論が証明されつつあり(Shidara, 1993)、仮想軌道制御仮説を否定する証拠を提出している(Gomi, 1996)。この理論の反射に対する考え方は明らかではないが、随意運動中は反射は抑制されているとみなしているようである。あるいは、反射は神経系のハードウエアの一部として、随意運動と同時に制御されるという立場ととらえることもできる。現在の筆者はこの立場に近く、反射がさまざまな運動の専用回路となっていることで脳の計算が省力化されているのではないか、と考えている。計算理論の今後の発展が期待される。
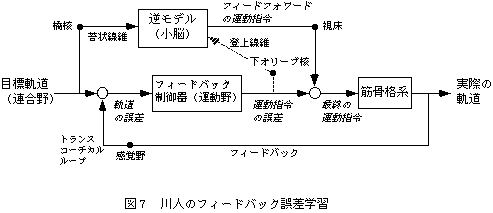
図5 川人のフィードバック誤差学習
フィードバック誤差学習では制御と学習が同時にすすむ。フィードバック情報は小脳の内部モデルを学習するために重要である。学習がすすむにつれ、フィードバック回路を流れる情報は減少し、小脳を中心としたフィードフォワード回路によって制御されるようになる。
9.おわりに
各解説にあるように、反射をめぐって観察される現象には、生理学的に興味深い現象が多く、随意運動制御全体が主に反射によって制御されているのではないかと感じるほどの驚きをもってデータをみることも多い。また、反射学の発展の背景には、病的反射や痙縮など、診断に必要な徴候であったり、そのものが治療の対象となっているという状況があり、実際、反射の異常が運動障害の原因となっている場合も多い。しかしこれは、正常の運動制御とは分けて考えるべきであろう。反射は一部であって全体ではない。基礎研究者であれ臨床家であれ、反射を過大評価も過小評価もすることなく、その役割と位置付けを今後明確にしていくことが重要であると考える。
文 献
Bernstein, NA: The co-ordination and regulation of movements. Pergamon Press. New York, 1967
Bernstein, NA: The problem of interrelation between coordination and localization. Arch Biol Sci, 38: 1-35 (in Russian), 1935
Bizzi E, Accornero N, Chapple W, Hogan N: Posture control and trajectory formation during arm movement. J Neuroscience 4: 2738-2744,1984
Domen K, Latash ML & Zatsiorsky VM: Reconstruction of equilibruim trajectories during whole-body movements. Biol Cybern 80: 195-204, 1999
Feigenberg IM & Latash LP: N. A. Bernstein: The Reformer of Neuroscience. In Dexterity and its development. Latash ML & Turvey MT (Eds.) pp247-275, Lawrence Erlbaum Mahwah NJ, 1996
Feldman AG, Adamovitch SV, Ostry DJ, Flanagan JR: The origin of electromyograms -Explanations based on the equilibrium point hypothesis. In JM Winters & SL-Y Woo (Eds.) Multiple muscle systems. Biomechanics and movement organization. pp. 195-213.Springer-Verlag, New York,1990
Feldman AG, Functional tuning of the nervous system with control of movement or maintenance of a steady posture II. Controllable parameters of the muscle. Biophysics 11: 565-578, 1966
Feldman AG, Functional tuning of the nervous system with control of movement or maintenance of a steady posture III. Mechanographic analysis of execution by man of the simplest motor task. Biophysics 11: 667-675, 1966
Feldman, AG: Once more on the equilibrium-point hypothesis (λmodel) for motor control. J Mot Behav 18: 17-54, 1986
Gomi H, Kawato M: Equilibrium-point control hypothesis examined by measured arm-stiffness during multi-joint movement. Science, 272: 117-120, 1996
Hoffer JA & Andreassen S: Regulation of soleus muscle stiffness in premammillary cats. Intrinsic and reflex components. J Neurophysiol 45: 267-285, 1981
Kawato M, Furukawa K, & Suzuki R. A hierarchical network model for motor control and learning of voluntary movement. Biol Cybern 57: 169-185, 1987
川人光男:脳の計算理論, 産業図書,東京,1996
小宮山:バイオメカニズム学会誌 : ,1999
Latash ML & Gottlieb GL: Virtual trajectories of single-joint movements performed under two basic strategies. Neurosci 47: 357-365, 1992
Latash ML: Control of Human Movement. Human Kinetics. Champaign IL. 1993
Latash ML: Neurophysiological Basis of Movement. Human Kinetics. Champaign IL. 1998
Merton PA: Speculations on the servo-control of movements. In : The spinal cord (ed. by JL Malcolm, JAB Gray & GEW Wolstenholm), pp. 183-198, Little Brown, Boston, 1953
Shidara M, Kawano K, Gomi H, KawatoM: Inverse-dynamics encoding of eye movement by Purkinje cells in the cerebellum. Nature 365: 50-52, 1993