Interactions Non-Locales dans l'Expression des Gènes
Joël Sternheimer
Université Européenne de la Recherche, 1 rue Descartes 75005 Paris
Résumé: La prise en compte des interactions non-locales dans le génôme et l'expression des gènes conduit à montrer l'existence et à préconiser l'emploi de méthodes spécifiques de régulation épigénétique -- c'est-à-dire agissant sur cette expression pour la stimuler ou l'inhiber --, généralement bien mieux adaptées à leur objet que l'utilisation d'OGM.
l. Comme l'avait pressenti le physicien Hermann Weyl en 1918 (1), l'invariance des lois physiques par rapport au choix de l'unité de mesure, loin d'être une propriété purement numérique ou géométrique ayant seulement trait à leur représentation, a bel et bien des conséquences physiques mesurables (2) - la mesure d'un objet par un observateur n'étant qu'un cas particulier de l'altérité, nécessairement réciproque, des objets physiques les uns relativement aux autres. L'invariance d'échelle de l'instrument de mesure (avec un rapport donné de chaque unité à la suivante), nécessaire pour pouvoir mettre en évidence cette propriété, s'étend ainsi aux objets en général pour autant et dans les limites où chacun peut effectivement être pris comme instrument de mesure par rapport à un autre.
Les ondes d'échelle
Une conséquence de cette invariance est alors l'existence d'ondes reliant entre elles des échelles différentes, et qui généralisent en ce sens les ondes associées aux particules par la mécanique quantique (2). Ceci entraîne que, dans un objet qui en est le siège, la précision relative d'une mesure est limitée en elle-même par la portée de l'échelle (3). En d'autres termes, si l'on réduit cette portée en décomposant l'objet afin d'en étudier les composants, on perd irréversiblement de l'information : si on "démonte" un objet qui est le siège d'ondes d'échelle, on ne peut plus le remonter. Mais inversement, lorsque cette dernière propriété est vérifiée, l'existence conséquente de ces ondes implique qu'il doit exister des traces à l'échelle de l'objet entier de ce qui se passe à d'autres échelles plus fines ; leur "lecture" permet alors non seulement d'étudier cet objet, mais aussi dans une certaine mesure d'interagir avec lui. Le respect de l'objet d'étude, qui découle de la nécessité de préserver la portée de l'échelle afin d'assurer aux mesures une précision suffisante, est alors, dans ce cas, l'attitude naturelle à adopter sur le plan méthodologique (3 et 4).
2. On aura reconnu dans la propriété d'impossibilité de remonter un objet une fois qu'on l'a démonté, une propriété que possèdent, notamment, les êtres vivants. Cela signifie que dans ce qui caractérise précisément ces êtres vivants, à savoir l'existence d'un génome et l'expression des gènes composant ce dernier lors du processus de synthèse des protéines, on peut s'attendre à ce que des ondes d'échelle se manifestent et soient décelables. Et qu'en conséquence soient présentes dans ce génome des interactions non-locales, impliquant différentes échelles de son organisation, et donc une limitation essentielle à toute entreprise de le modifier localement; mais aussi la possibilité d'agir sur son expression de façon spécifique, grâce aux lois qui y président.
Des ondes dans l'expression des gènes
Considérons en effet ce qui se produit lors de l'expression des gènes (5) . Lorsqu'on absorbe des aliments, le processus de digestion qui s'ensuit consiste à les décomposer en éléments simples, les sucres en oses, les graisses en acides gras et les protéines en acides aminés. Notre programme génétique, contenu dans notre ADN, nous permet ensuite de fabriquer nos propres protéines à partir des acides aminés provenant de l'alimentation ou - pour certains - produits par l'organisme lui-même. De l'ADN va se détacher un ARN messager, qui va lui-même se fixer sur le ribosome, endroit très stable dont la forme d'établi va permettre la fixation des molécules nécessaires à la synthèse des protéines. Puis les ARN de transfert (tARN), qui portent les acides aminés, vont à leur tour se fixer sur l'ARN messager déjà fixé sur le ribosome. Par déplacements successifs, les acides aminés portés par les tARN vont alors s'accrocher les uns aux autres pour former une chaîne protéique : un acide aminé, situé au bout d'un tARN, va s'accrocher à un autre acide aminé, bientôt suivi d'un deuxième, puis d'un troisième, et ainsi de suite.
Si ce qui précède est aujourd'hui bien connu, considérons maintenant plus précisément ce qui se passe (6) au moment où l'acide aminé, porté par son tARN, vient s'accrocher sur le ribosome. Car lorsque l'acide aminé est à l'état libre, son comportement est simplement celui d'une particule soumise à l'agitation thermique : sa longueur d'onde de de Broglie est alors négligeable au regard de sa taille. Mais lorsqu'il s'accroche sur le tARN, ce n'est plus tout à fait le cas: fixé par un seul degré de liberté, il conserve une relative autonomie, tout en étant déjà fortement ralenti vis-à-vis de l'agitation thermique. Sa longueur d'onde, inversement proportionnelle au produit de sa masse par sa vitesse, cesse alors d'être négligeable et ses propriétés quantiques commencent à apparaître. Et lorsque le tARN qui le porte vient se fixer sur le ribosome, la stabilisation vis-à-vis de l'agitation thermique devient considérable, et la longueur d'onde de l'acide aminé dépasse sa taille de plusieurs ordres de grandeur : son comportement est alors celui d'une onde, ce qui signifie que, pendant le bref instant où cela se produit, l'acide aminé émet un signal.
Bien sûr, immédiatement après, le comportement de l'acide aminé redevient celui d'une particule : du point de vue de la mécanique quantique ordinaire (7) , si l'acide aminé est, pour parler simplement, particule puis onde puis particule, cela veut dire que le signal a disparu dès que le comportement corpusculaire réapparaît. En fait, c'est là que la considération selon laquelle, dans ce système intégré à plusieurs échelles, la fonction d'onde quantique doit être généralisée en une fonction d'onde d'échelle permet une prédiction originale. Selon celle-ci en effet, à la particule est associée une onde à valeurs non seulement dans l'espace-temps mais dans l'échelle, dans laquelle elle se propage comme une somme d'ondes de vitesses différentes, passant d'une échelle plus petite - celle de l'acide aminé - à une plus grande - celle de la protéine en formation - en des temps multiples entiers de celui mis par l'onde la plus rapide. De là vont nécessairement résulter des corrélations temporelles entre les fréquences associées aux acides aminés séparés par un multiple entier d'une certaine période. D'où une vérification simple : ces corrélations sont-elles ou non observées ?
Des effets non-locaux vérifiables expérimentalement
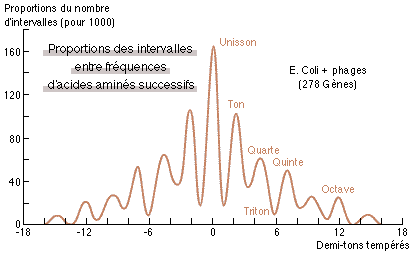
3. La réponse est oui, et plus encore. Considérons en effet la suite des fréquences associées à la synthèse d'une protéine donnée quelconque. Si, dès la fixation de chaque acide aminé sur l'ARN de transfert, les propriétés ondulatoires se manifestent déjà, il va en résulter une première prédiction: celle de la synchronisation sur harmoniques des fréquences de cette protéine, au bout d'un temps très court (de l'ordre de l0-12,5 seconde) - de la même façon que les masses des particules élémentaires les plus stables (précisément plus stables que cette durée) se synchronisent sur une même gamme musicale (8). Le résultat d'une telle synchronisation peut alors se calculer aisément pour une proportion donnée de tARN dans une cellule (cf.(9)). Dans ces conditions, les corrélations temporelles mentionnées ci-dessus vont entraîner à leur tour une prédiction : à savoir que les intervalles de fréquences associées aux acides aminés successifs vont tendre eux aussi, à se synchroniser sur les harmoniques de rang le plus bas possible, autrement dit favoriser les intervalles consonants (au sens de Hellegouarch (l0)) par rapport aux dissonants. La figure 1 ci-dessous montre ce qu'il en est pour 278 gènes procaryotes (colibacille et phages) (11) ; montrant très clairement (cf. aussi fig.2) l'existence des interactions non-locales prédites.
| Fig.1: Lissage des proportions pour 1000 du nombre d'intervalles entre fréquences d'acides aminés successifs, pour 278 gènes procaryotes; en abscisse, les intervalles (rapports de fréquences) en demi-tons tempérés (puissances de 2 1/12). La position des maxima et minima, correspondant à des intervalles respectivement consonants et dissonants, révèle la présence (et l'amplitude relative) d'harmoniques dans les ondes associées aux acides aminés individuels, ainsi que leur relative persistence d'un acide aminé à l'autre lors du processus de synthèse. |
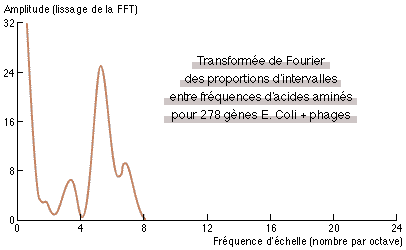
| Fig.2: Transformée de Fourier rapide (FFT) du signal de la figure 1. Le pic observé (à la fréquence d'environ 5 par octave) constitue en lui-même une signature de la présence d'interactions non-locales dans l'expression des gènes. |
Si l'on prend plus précisement comme indice le rapport unisson (fréquence identique) sur triton (quarte augmentée, particulièrement dissonante) on observe (fig.3) une croissance régulière de cet indice suivant que l'on considère, respectivement : des séquences au hasard, correspondant <1> à des fréquences équiprobables (c'est à dire réparties en proportions égales), <2> à des acides aminés équiprobables, <3> à des codons équiprobables, <4> à des acides aminés selon leurs proportions observées (12) ; puis à des séquences réelles, <5> chez des procaryotes (11) et <6> chez l'homme (séquence de la titine, la plus grosse protéine connue, comprenant 26 926 acides aminés (13)).
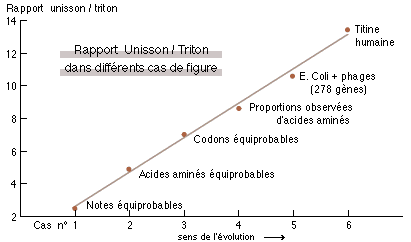
| Fig. 3: Variation du rapport des proportions d'intervalles d'unisson (fréquences successives identiques) sur triton (quarte augmentée) pour des séquences au hasard construites à partir de notes équiprobables, d'acides aminés équiprobables, de codons équiprobables, et des proportions observées d'acides aminés ainsi que pour les séquences réelles des 278 gènes procaryotes de la fig. 1, et des 26926 acides aminés de la titine humaine. |
En outre les "périodes" prédites sont bien observées, et sont, tout comme les répartitions d'intervalles ci-dessus, caractéristiques d'une protéine donnée (cf.(9)). Mais surtout une propriété essentielle apparaît : les protéines ayant des suites de fréquences homologues sont de façon systématique, métaboliquement agonistes, montrant que ces suites de fréquences ont une activité métabolique de régulation épigénétique réciproque des biosynthèses protéiques. En effet, les amplitudes des "ondes d'échelle" associées à des séquences identiques s'ajoutant, vont correspondre par leur carré à la probabilité que ces séquences soient exprimées dans l'organisme considéré : les fréquences identiques vont ainsi interagir les unes sur les autres par un phénomène de type résonance, et la synthèse d'une protéine va rétroagir positivement sur les autres protéines de l'organisme ayant des suites de fréquences homologues.
Considérons par exemple le cytochrome C, avant-dernière protéine de la chaîne respiratoire. Sa période est de 8 acides aminés, ce qui correspond à un "thème" caractéristique (9) . Dans la dernière protéine de cette chaîne, la cytochrome oxydase, on retrouve effectivement la présence de ce "thème", signifiant que sa synthèse va, en elle-même, "relancer" dans l'organisme concerné la synthèse du cytochrome C, ce dont l'intérêt régulateur pour l'organisme est évidemment manifeste. Il est à noter qu'il s'agit là de suites de fréquences identiques, alors même que les séquences d'acides aminés et donc encore plus de nucléotides correspondants diffèrent : il s'agit donc bien d'une régulation non-locale, distincte d'une interaction chimique ordinaire.
La généralité de ces propriétés peut alors être vérifiée grâce au moyen que nous avons ainsi de prédire les fonctions des protéines à partir de leur séquence d'acides aminés. Et en effet, elle se vérifie de façon systématique (9 et 14). De plus, la transposition à notre échelle de ces suites de fréquences a une action - par résonance d'échelle - sur la synthèse de la protéine correspondante, permettant de réguler épigénétiquement l'expression des gènes, de manière beaucoup plus souple et contrôlable qu'une modification du génome.
Un certain nombre d'expériences ont ainsi été conduites, où en agissant sur l'expression d'un gène donné (soit pour la stimuler comme ci-dessus, soit pour l'inhiber avec des suites de fréquences symétriques (9)), les conséquences à notre échelle en sont systématiquement observées - ce qui, en respectant l'objet d'étude, permet un suivi particulièrement fin et détaillé (15 et 16) .
Nous disposons donc d'un moyen pour stimuler ou inhiber fonctionnellement et réversiblement l'expression d'un gène donné, d'un facteur pouvant aller jusqu'à 20 environ, permettant de répondre avec une bien meilleure finesse et souplesse d'adaptation à beaucoup de problèmes pour lequel le recours aux OGM est aujourd'hui envisagé.
Des actions qui - d'abord - respectent leur objet
4. Considérons en effet les implications de ce qui précède au regard de ce qui constitue l'objet de cet ouvrage, les organismes génétiquement modifiés. Le principe de ces modifications génétiques consiste à insérer un gène extrait d'un génome dans un autre. Il est clair qu'en général ce ne sera tout simplement pas possible, du fait des nombreuses interactions non-locales qui, ainsi que nous l'avons vu, régulent ce génome. Cependant il est des cas où cela sera possible, mais alors avec pour conséquence des modifications du génome résultant, pour le rendre compatible avec les lois qui gouvernent ces interactions : l'organisme résultant va évoluer. Le fait qu'il ne suffit pas que l'insertion ait lieu pour garantir la stabilité et l'innocuité du résultat est attesté, pour ne prendre qu'un exemple, par l'existence de rétrovirus pathogènes. Le maïs transgénique Bt est, de son côté, basé sur un postulat qui découle directement d'une image "mécanique" du vivant : pour qu'un maïs ne soit pas attaqué par les insectes prédateurs, lui insérer un gène insecticide... avec la première conséquence d'un maïs - et son environnement (17) - rendus toxiques à la longue; la deuxième du développement (en à peine quelques années (18)) d'une résistance chez les insectes visés ; la troisième, d'un risque de perturber le cycle de croissance de la plante, du fait d'homologies de suites de fréquences (du type de celles signalées plus haut (14)), entre des passages du transgène Bt et de protéines de division cellulaire du maïs; et enfin... d'être simplement passé à côté de la question. En effet, la prise en compte des lois d'interactions non-locales mentionnées ne fournit pas seulement la possibilité d'un contrôle ou d'une sélection au cas par cas, que peut déjà permettre la considération des homologies que nous venons d'évoquer : elle amène surtout, dans la plupart des cas, à considérer qu'il y a mieux à faire que de modifier génétiquement un organisme pour résoudre un problème. Ainsi de l'expérience réalisée en Afrique (M. Gueye et al.(l6)) sur des tomates rendues épigénétiquement résistantes à la sécheresse : elles étaient également épargnées par les insectes prédateurs qui préféraient s'en prendre aux plants témoins moins vigoureux qu'aux plants sur lesquels avait eu lieu l'expérience; simplement (comme certes il conviendrait, en la matière, de ne pas l'oublier) parce que la tendance naturelle de ces prédateurs est quand même d'abord de respecter la nature dans laquelle ils vivent, aussi s'en prennent-ils de préférence aux plants les moins vigoureux, épargnant les autres. Il est évident qu'ils trouvent là un avantage évolutif, puisqu'ils contribuent ainsi à préserver la planète sur laquelle ils vivent.
Références
(1) H. Weyl, "Raum Zeit Materie", Berlin (l9l8); trad. angl. "Space time matter", Dover ed., p. 282 (1952).
(2) J. Sternheimer, exposé au Colloque international "Louis de Broglie, Physicien et penseur", Anc. Ecole Polytechnique, Paris, 5-6 novembre 1987;
"Ondes d'échelle : une expression spécifique de la covariance des lois physiques lors d'un changement d'échelle d'observation", soumis aux C. R. Acad. Sc. Paris (1997).
(3) J. Sternheimer, Le Cahier du Collège international de philosophie n° 3, pp. l80-l82, Osiris, Paris 1987.
(4) G. Huber, in Psychanalyser après la choa, pp. 147-153, Osiris, Paris 1988; V. Bargoin, "Après le séquençage de l'ADN du génome de la levure : le face-à-face entre la science et l'éthique", Le quotidien du médecin n° 6089, p. 10, 18 juin 1997.
(5) Cf. par exemple B. Alberts et al., Biologie moléculaire de la cellule, chapitre 6, Flammarion, Paris (3ème édition1995).
(6) J. Sternheimer, "Epigenetic regulation of protein biosynthesis by scale resonance", exposé à l'Académie des Sciences de Tokyo-Kanagawa (23/5/1993), repr. partielle in Science Frontières n° 7, p. 6 (avril 1996).
(7) Cf. par exemple l'exposé d'A. Aspect au Colloque international "Louis de Broglie, physicien et penseur", loc. cit. (1987).
(8) J. Sternheimer, C. R. Acad. Sc. Paris, vol. 287, pp. 829-834 (1983); séminaire de physique mathématique, Collège de France (31/1/1984), repr. in Rev. Bio-Math., n° 94, pp. 1-47 (1986).
(9) J. Sternheimer, Procédé de régulation épigénétique de la biosynthèse des protéines par résonance d'échelle, brevet n° FR 92 06765 (1992), délivré le 13/7/95.
(10) Y. Hellegouarch, C. R. Math. Rep. Acad. Sci. Canada, vol. 4, pp. 277-281 (1982).
(11) Cf. A. S. Kolaskar et B. V. B. Reddy, J. Biomol. Struct. Dyn. vol. 3, pp. 725-738 (1986).
(12) M. D. Dayhoff, Atlas of protein sequence and structure, vol. 5, Suppl. 3, chap. 25, N.B.R.F. (Washington) 1978.
(13) S. Labeit et B. Kolmerer, Science vol. 270, pp. 293-296, 1995.
(14) J. Sternheimer, "Quelques prédictions des homologies musicales sur les fonctions épigénétiques des protéines", in réponse à la N.O. OEB 93 913 082.9 du 28/0l/97 (23/05/97).
(15) P. Ferrandiz, "Procédé de régulation épigénétique de la synthèse protéique : essais en panification", Industries des Céréales n° 85, pp. 40-42 (l993);
"De la musique et des plantes", La Garance Voyageuse n° 37, pp. 25-28 (1997).
(16) M. Ulmer, B. Gil, P. Ferrandiz et J. Sternheimer, "Régulation
épigénétique de la biosynthèse des protéines
appliquée à la culture de fruits et légumes : compte-rendu
d'expérience en jardin potager" (1993),
cf. J.-M. Pelt, "Les langages secrets de la nature", chap. XVIII,
Fayard, Paris1996;
S. & S. Saito et J. Sternheimer, "Effet de la stimulation épigénétique de la chalcone isomérase sur la coloration des pommes" (1993);
J.M. Huber, J. F. Treyvaud, B. Dubouloz, C. et R. Egloff, A. Lappert
et J. Sternheimer, "Régulation
épigénétique de la biosynthèse des protéines
appliquée à la culture de tomates : compte-rendu d'expérience
en serre" (1994),
cf. Science Frontières n° 7, pp. 2-7 (avril 1996);
P. Ferrandiz, "Stimulation épigénétique de protéines de phototosynthèse sur culture d'algues bleues cyanophycées : un modèle pour la réduction de la pollution atmosphérique" (1995);
M. et O. Gueye, F. Diagne, J-J. Houziel, P. Ferrandiz et J. Sternheimer,
"Stimulation épigénétique
de la résistance à la sécheresse pour des cultures
de tomates: une expérience en plein air au Sénégal",
rapport UER (1996),
cf. Science Frontières n° 14, p. 3, décembre1996;
F. Sneyaert, P. Ferrandiz, M. Renoma et J. Sternheimer, "Conservation de fruits et légumes par régulation épigénétique : inhibition de l'expression de la polygalacturonase d'avocat" (1997), in réponses aux N.O. OEB du l8.08.95, 23.01.97 et 11.07.97 (l996-1997).
(17) Cf. C. J. Palm et al., Mol. Ecol. vol. 3, pp. 145-l5l (1994) ;
G. E. Séralini, communication au Colloque OGM organisé par l'Université Européenne de la Recherche et Ecoropa au Carré des Sciences, Paris (21 avril 1997).
(18) B. E. Tabashnik, Proc. Nat. Acad. Sci. USA vol. 94, pp. 3488-3490 F. Gould et al., id., pp. 3519-3523, april 1997.
![]()