運動制御 Motor Control
(現代リハビリテーション医学 金原出版より P41~46) 道免和久
運動障害を治療するハビリテーション医療において、正常の随意運動制御の理解は不可欠である。運動制御の諸理論には、運動障害の理解のみならず運動学習など運動療法の基礎の理解のためにも重要な概念を含んでいる。
運動制御とエングラム
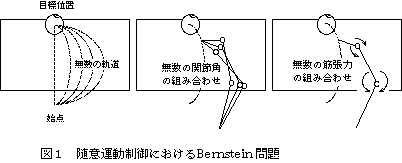
人間がテーブルの上のリンゴを取る動作を考えるとき(図1)、リンゴまでの軌道は無数に存在しうるが、自然な随意運動では最適な1つの軌道が選択される。軌道が決まっても、関節角の組み合わせは無数に存在しうるが、これも最適な1つの組み合わせが選択される。さらに、関節角が決まっても、筋肉の張力の組み合わせは無数に存在するはずであるが、ここでもある1つの組み合わせが選択される。このように随意運動では、多くの自由度を有する筋骨格系が、中枢神経によって見事にあやつられ、なめらかで、かつやわらかい運動が実現されているのである。このような随意運動の特徴は、1930年にBernsteinによって指摘されているため、Bernstein問題[1]とも呼ばれる。Bernsteinによれば、Bernstein問題を解決するために中枢神経は、作用の似たいくつかの筋群が共同して働くシナジー(synergy)を利用しているといわれている。
シナジーの存在は、複雑な筋骨格系を制御しやすくしているが、実際の随意運動は、さらに多様性に富み、かつ正確である。もし、一つ一つの筋肉への運動指令が脳の中で別々にスイッチされている(図2)とすると、多関節の多様な運動を瞬時に行うことはできないであろう。そこでBernstein[2]は、運動全体の抽象的な形での記憶が脳に蓄えられていると考え、これをエングラムと呼んだ。シナジーのもとにもなる運動プランと言ってもよい。Kottke[3]はこの概念をリハビリテーションに適用し、多くの筋群への一連の運動指令パターンをエングラムと定義した。Kottkeは、あらかじめプログラムされたエングラムが適切なトリガーに反応して発現するようにエングラムの強化をすることによって、運動の協調が可能になると考えた。エングラムの強化には、数多くの運動の繰り返しが必要であるが、いったんエングラムが確立されれば、複雑な随意運動が協調性をもって実行されることになる。このような概念的理解は、徐々に神経科学を中心に実証されつつある。
フィードバック制御
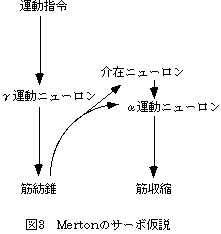
遅い簡単な運動は、体性感覚や視覚のフィードバックによって遂行可能である。また、脊髄レベルでのフィードバックループを前提とした制御理論としてサーボ仮説[4](図3)があり、γ運動ニューロンに運動指令が与えられることによって、反射的にα運動ニューロンが活動して随意運動が起こるといわれている。しかしいずれの制御でも、フィードバック時間の遅れなどの理由により、速い運動を制御することは理論的に不可能とされている。
仮想軌道制御仮説
フィードバックによらないフィードフォワードの運動を説明するために、仮想軌道制御仮説[5]が提案されている。これは、運動指令が筋肉のばね特性を調節して、その釣り合い位置に向けて関節が動く、という仮説である。つまり随意運動は、経時的に変化する釣り合い位置(仮想軌道)に関節が追従することによって発現するという(図4)。仮想軌道制御仮説には、α運動ニューロンの活性を調節しているという仮説(αモデル)と、緊張性伸張反射の閾値を調節して筋長を制御しているという仮説(λモデル)[6]がある。
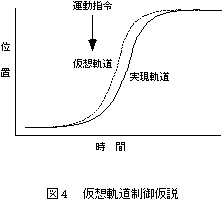
仮想軌道制御仮説は、中枢神経が複雑な動力学的計算をせずに、軌道を直接調節できるという点で魅力的であるが、実現軌道に似た鋳型のような軌道にわずかに遅れて筋骨格系が追従できるためには、剛性(スティフネス)が高くなければならない。つまり「かたい」制御を行っているという前提が必要となる。ところが、実際に運動中のスティフネスは予測より低く、仮想軌道も単純な形ではないことが実証されてから[7]、仮想軌道制御仮説はその論拠を失いつつある。
内部モデル制御と学習モデル
小脳皮質は層状の均一な構造をしており、学習が可能な神経回路(ニューラルネットワーク)の集まりであると考えられている。小脳が学習機能をもつことは、伊藤ら[8]によるプルキンエ細胞の長期減弱の研究等から知られている。
運動中の学習によって小脳は、筋骨格系への入出力関係、つまり、運動指令とその結果生じる軌道との関係の情報を蓄える[9,10]。このような小脳内に保持される運動指令-筋骨格系の情報は内部モデルと呼ばれ、運動指令から軌道を出力する神経回路を順モデル、逆に軌道に見合った運動指令を出力する神経回路を逆モデルと呼ぶ。
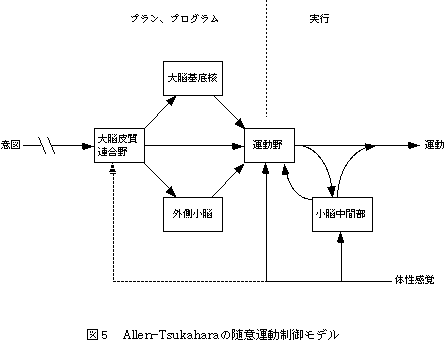
Allen-Tsukahara[11]の随意運動の制御モデルによれば(図5)、大脳皮質連合野が小脳外側部と大脳基底核の助けを借りて運動が計画された後、運動野から運動指令が脊髄を通して筋骨格系に送られる。体性感覚情報は、小脳傍虫部にフィードバックされ運動の修正を行う。
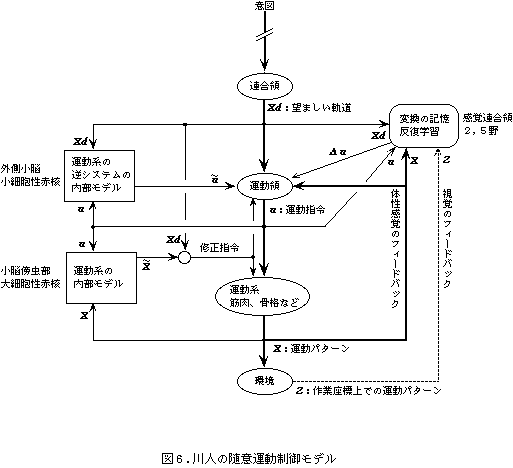
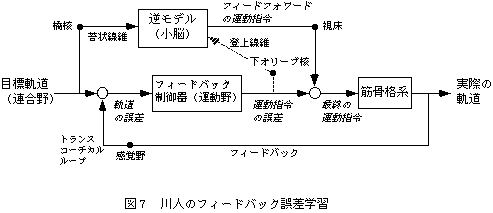
川人は、Allen-Tsukaharaの制御モデルを、神経回路の理論を応用して発展させ、以下のモデルを提案した(図6、7)。まず連合野から運動野に目標軌道が送られ、運動野から運動指令が脊髄へ伝えられる。実現した運動の情報は、大脳皮質を介するトランスコーチカルループによって運動野にフィードバックされる。このフィードバック回路でも運動は可能だが、フィードバック時間の遅れなどの理由で、速い滑らかな運動はできない。そこで、小脳外側部-赤核系は、目標軌道と運動指令をモニターし、運動にみあった運動指令を出力する内部モデル(逆モデル)を学習によって小脳に形成する。学習がすすめば速いスムーズな運動がこの回路を用いて可能になる。小脳傍虫部-赤核系は、実現軌道のフィードバック情報と運動指令を受け取ることで、運動指令から軌道を予測する内部モデル(順モデル)を小脳に形成するので、運動前に軌道誤差を予測して運動指令の修正を行うことが可能になる。目標軌道と実現軌道の間の誤差は、誤差信号(教師信号)として下オリーブ核から小脳に伝達され、小脳の内部モデルの修正が行われる。これをフィードバック誤差学習といい、運動制御と運動学習を統合した理論といえる。
運動学習とエングラム
Kottke[12]は、運動学習においてエングラムの強化が重要であるという仮説を提案してきたが、脳研究の進歩によりエングラム強化のメカニズムの本態が次第に明らかになってきた。
運動学習は、movement(動き)の学習とprocedure(手続き)の学習に分けられる[13]。movementの学習については、内部モデルを含む制御モデルで述べた通り、小脳を中心に、大脳皮質と小脳との相互作用で行われる。この場合、小脳において目標軌道が各筋群への運動指令に変換されるという点で、エングラムは小脳に蓄えられていると考えることができる。
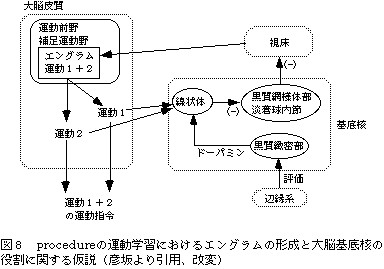
一方、一定の順序をもった多くの要素からなる運動では、そのprocedureを学習する必要がある(図8)。procedureの学習は大脳基底核と大脳皮質との相互作用で行われている、という仮説が提案されている。すなわち、連続した運動結果の情報は線状体に集まり、辺縁系がこの結果を'快い'ものと評価すると、視床を介して大脳皮質のprocedureをコードしたニューロン群の活動が高まると考えられる。学習を繰り返すと、運動のprocedureが補足運動野や運動前野に蓄えられる。これはKottkeの指摘したエングラムに相当すると考えられる。procedureの学習の中でも、運動要素個々の協調性はやはりmovementの学習として小脳によって獲得される。
運動関連領野と基底核の役割
情動、モチベーションに関与する帯状回の活動は、自発性の運動発現にも重要である。補足運動野は、帯状回の入力を強く受け、複雑な運動を行うときだけでなく、運動をイメージするだけでも活動することから、運動のプログラミングに関与すると考えられている。運動前野は、特定の動作すなわち運動のサブプログラムをコードしているといわれ、頭頂連合野→運動前野→一次運動野の経路が、外界の認識→動作への変換→運動のサブプログラム形成→運動指令の過程に対応すると考えられている[14]。
基底核出力部(淡蒼球内節、黒質網様体部)は、視床、脳幹部等を持続的に抑制しており、線状体(尾状核、被殻)の活動による脱抑制、あるいは、線状体から淡蒼球外節、視床下核を介する抑制強化により、運動を選択する役割をもつ。すなわち、基底核は、大脳皮質からの入力を受けながら、視床を介して運動前野、前頭連合野などをコントロールするというループ回路を形成している。さらに、基底核には辺縁系からの強い入力があり、これが行動の評価(価値判断)と行動の学習に役立っているといわれている[13]。
姿勢制御の仕組み
姿勢制御は、外乱に対して姿勢を維持するための姿勢反射と、上肢など身体の一部の随意運動を達成するために無意識的に調節される姿勢調節に分けられる[15]。
1) 姿勢反射
人間の立位姿勢は、逆さ振り子inverted pendulumに例えられるように、高い位置にあって不安定な身体重心のずれを反射等を利用して修正することによって維持される。脳幹からの網様体脊髄路によって伸筋が持続的に活動することで立位が保持されるが、重心の移動は脳によって常にモニターされている。外乱に対しては、下肢遠位の関節から頚部の順に反射によって姿勢の修正が行われ、重心の乱れが抑えられる。
姿勢反射は、長潜時の伸張反射stretch reflex、脊髄での伸張反射、支持反応supporting reactionなどの局所性姿勢反応、交叉伸展反射crossed extension reflex、屈曲反射flexion reflexなどの体節性姿勢反応、緊張性頚反射tonic neck reflex、立ち直り反射reghting reflexなどの全身性姿勢反応に分類される。これらの反射や反応は、脊髄、延髄、中脳など各部位が関与し、大脳基底核によって統合される。小脳虫部には、体幹の状態がフィードバックされ、姿勢を調節しながら、体幹の内部モデルが形成されることで、環境や運動に応じた調節が可能になるといわれている[10]。
2) 随意運動遂行時の姿勢制御
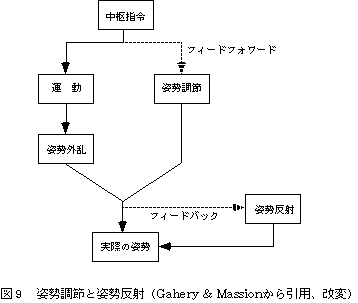
ある姿勢をとった状態で随意運動遂行するとき、たとえば、立位において上肢を挙上するとき、上肢の筋電図に先行して腓腹筋の活動が生じる。この活動は、腕の運動に備えて予測的に姿勢が調節されていることを意味している。姿勢調節は腓腹筋のほか、ハムストリングス、傍脊柱筋にも生じ、遠位筋から近位筋の順に活動することが知られている。このように随意運動に先行する姿勢調節はanticipatory postural adjustment[16]とよばれる。すなわち、随意運動によって姿勢の乱れが生じることがあらかじめ予測できる状況では、フィードバック機構による姿勢反射がはたらく以前にフィードフォワード的に姿勢調節がなされる[17](図9)。
Nashner[18]は随意運動に平行した姿勢調節をpostural setとよび、大脳連合野、大脳基底核、新小脳によって運動指令が準備されると同時に、最適な姿勢が選択されるといわれている。
歩行制御の仕組み
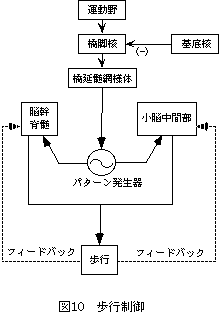
歩行運動は一定のパターンの運動の集まり、つまりシナジーであると考えられている[19]。動物実験の結果から、歩行のパターンにおけるリズム生成および肢間協調の中枢が脊髄にあることがわかっており、中枢パターン発生器central pattern generator[20]とよばれている(図10)。中脳の歩行誘発野mesencephalic locomotor region(=橋脚核pedunculs pontine nucleus)からの持続的な入力により、脊髄の中枢パターン発生器が歩行リズムを生成する。歩行誘発野には基底核群、感覚運動皮質、辺縁系からの入力があり、橋延髄網様体と脊髄介在ニューロンに出力を送る。脊髄の中枢パターン発生器では、屈筋と伸筋の脊髄介在ニューロン群相互にある抑制性結合が屈曲伸展の交互運動を生み出し,複数の脊髄介在ニューロンの回路網の興奮が循環することによって律動的発振を発生すると考えられている。末梢からの入力のうち、Golgi腱器官からのIb群感覚信号は、中枢パターン発生器に対して強く影響する[21]。小脳中間部は、歩行誘発野からの情報と四肢からのフィードバック情報の両方をうけとり、歩行に関する内部モデルを形成していると考えられている[10]。
[文 献]
1) Bernstein, NA :The co-ordination and regulation of movements. Pergamon Press. New York, 1967
2) Bernstein, NA: The problem of interrelation between coordination and localization. Arch Biol Sci, 38: 1-35 (in Russian), 1935
3) Kottke FJ, Halpern D, Easton JKM, Ozel AT, and Burrill CA: Training of Coordination. Arch Phys Med Rehabil 59: 567-572,1978
4) Merton PA: Speculations on the servo-control of movements. In : The spinal cord (ed. by JL Malcolm, JAB Gray & GEW Wolstenholm), pp. 183-198, Little Brown, Boston, 1953
5) Bizzi E, Accornero N, Chapple W, Hogan N: Posture control and trajectory formation during arm movement. J Neuroscience 4: 2738-2744,1984
6) Feldman, AG: Once more on the equilibrium-point hypothesis (λmodel) for motor control. J Mot Behav 18: 17-54, 1986
7) Gomi H, Kawato M: Equilibrium-point control hypothesis examined by measured arm-stiffness during multi-joint movement. Science, 272: 117-120, 1996
8) Ito M, Sakurai M, Tongroach P: Climbing fibre induced depression of both mossy fibre responsiveness and glutamate sensitivity of cerebellar Purkinje cells. J Physiol London 324: 113-134,1982
9) Kawato M, Furukawa K, & Suzuki R. A hierarchical network model for motor control and learning of voluntary movement. Biol Cybern 57: 169-185, 1987
10) 川人光男:脳の計算理論, 産業図書,東京,1996
11) Allen GI & Tsukahara N: Cerebrocerebellar communication systems. Physiol Rev 54: 957-1006, 1974
12) Kottke FJ: Therapeutic Exercise to Develop Neuromuscular coordination . In Krusen's Handbook of Physical Medicine & Rehabilitation (ed. by Kottke FJ & Lehmann JF), pp. 452-479, Saunders, Philadelphia, 1990
13) 彦坂興秀:随意運動における大脳基底核の役割. 脳とニューラルネット(甘利俊一、酒田英夫編),pp 218-234, 朝倉書店, 1994
14) 丹治順:運動関連領域野と運動プログラミング. 脳とニューラルネット(甘利俊一、酒田英夫編),pp 203-217, 朝倉書店,1994
15) 有働正夫:脊髄反射系と姿勢反射.日本臨床 45: 239-244, 1987
16) Cordo PJ & Nashner LM: Properties of postural adjustments associated with rapid arm movements. J Neurophysiol 47: 287-302. 1982
17) Gahery Y & Massion J : Co-ordination between posture and movement. Trends Neurosci. 199-202, 1981
18) Nashner LM: Adapting reflexes controlling human posture. Exp Brain Res 26: 59-72, 1976
19) 森茂美:歩行とその制御. 日本臨床45: 333-338, 1987
20) Grillner S: Control of locomotion in bipedes, tetrapods, and fish. In: Handbook of physiology- The Nervous System II ( ed by Brooks, VB) Waverly Press, Baltimore, 1981
21) 伊藤文男:筋感覚からみた運動制御、名古屋大学出版会、名古屋、1985
兵庫医科大学リハビリテーション医学
〒663-8501 兵庫県西宮市武庫川町1-1
Tel0798-45-6881
Fax0798-45-6948
E-mail: domen@neuro-reha.org